




合作客户/
拜耳公司 |
同济大学 |
联合大学 |
美国保洁 |
美国强生 |
瑞士罗氏 |






相关新闻Info
推荐新闻Info
-
> Kibron DeltaPi、Epsilon、Delta-8、Delta-12机型怎么选型
> 植物油、食用油界面张力,Kibron 检测油脂氧化变质程度
> 酸奶乳酸菌发酵液,Kibron 张力表征乳蛋白界面稳定性
> 食品饮料乳化剂 CMC,Kibron 高通量筛选食品级表活
> 天然植物护肤提取物表面活性,Kibron 超微量检测方法
> 牙膏润湿剂、发泡剂,Kibron 批量测试不同表活搭配效果
> 彩妆粉底液分散体系,Kibron 张力减少粉体沉淀分层
> 洗发水泡沫性能,通过 Kibron 动态张力评价起泡稳泡能力
> 天然植物精油防腐乳化体系,Kibron 张力优化复配比例
> 面霜乳液油水界面张力,Kibron 预判护肤品分层稳定性
不同种类的抗菌肽瓜娃素与生物膜之间的相互作用的性能对比【上】
来源:上海谓载 浏览 2132 次 发布时间:2022-07-05
摘要
颞叶素是最早从欧洲红蛙(Rana temporaria)的皮肤中分离出的短(10-13个氨基酸)线性抗菌肽,对革兰氏阳性细菌和白色念珠菌有效。为了深入了解其作用机制,我们比较了该家族的两个成员对模型膜的影响,即。,temporin B(LLPIVGNLLKSLL-NH2)和temporin L(FVQWFSKFLGRIL-NH2)。更具体地说,我们测量了它们插入脂质单分子膜以及它们对脂质体双层结构动力学的影响,如二苯基己三烯(DPH)和芘标记的磷脂所揭示的。我们还观察了这些肽对巨大囊泡拓扑结构的影响。这两种时间蛋白都很容易渗透到脂质单层中,在常见细菌带负电的磷脂-磷脂酰甘油的存在下,它们的嵌入作用增强。相反,真核细胞脂质胆固醇确实在一定程度上抵消了它们渗透到脂质膜中的作用。temporin B和temporin L均导致磷脂在双层中富集,并且在1-棕榈酰-2-油酰基-sn-甘油-3-磷酸甘油(POPG)存在下,这些肽增加了酰基链顺序。Temporin B实际上对由1-硬脂酰-2-油酰基-sn-甘油-3-磷酸胆碱(SOPC)组成的巨大脂质体没有影响,而当存在POPG时观察到快速发泡。相反,temporin L诱导SOPC和SOPC/POPG巨囊泡形成,而SOPC巨囊泡中胆固醇的存在减弱了这种作用。
抗菌肽广泛分布于自然界,是无脊椎动物和脊椎动物天然免疫的重要组成部分,可防止多种有害微生物的入侵和增殖。尤其是无尾类动物(青蛙和蟾蜍)的皮肤是抗菌肽的丰富来源,其结构和活性谱都具有很大的多样性(1,2)。通常大量生产,几种具有不同抗菌特性的不同肽通常存在于单个动物的皮肤提取物中。这种多样性被认为对保护青蛙免受更广泛的有害入侵者的侵害很重要(3)。蛙皮抗菌肽是在称为颗粒腺的特殊皮肤结构中产生和储存的,在肾上腺素能刺激或皮肤损伤时释放其内容物。一大类不断扩大的蛙抗菌肽,统称为前表皮抑制素,已被证明具有高度保守的N端前序列和具有与抗菌肽对应的可变序列的C端结构域(4)。
目前,对蛙皮和其他来源的抗菌肽进行了深入研究,以阐明其作用机制。作为这项工作的结果,抗菌肽通常被认为通过渗透或破坏细胞膜的稳定性来杀死靶细胞,无论是原核细胞还是真核细胞。然而,精确的机制仍不完全清楚。关于抗菌肽与生物膜和模型膜的相互作用,最近有许多优秀的综述,提供了迄今为止提出的用于解释肽诱导的膜渗透过程的不同模型的完整视图(5-8)。
颞蛋白是一类抗菌肽,首先从欧洲红蛙的皮肤中分离出来(9)。最近,在林蛙属其他物种的皮肤提取物中发现了颞蛋白家族的新成员,即。,R、clamitans(10)、luteiVentris(11)、pipiens(11)和grylio(12)。因此,这些发现也可以深入了解这些肽在相关物种之间的进化和分化(10,11)。颞蛋白是具有净正电荷和酰胺化C端的线性10-13残基长肽,在非极性溶剂(如三氟乙醇)中可能呈现两亲R螺旋构象(9,13)。发现时间蛋白A和B对大肠杆菌细胞壁缺陷突变株也有活性,并释放脂质体包埋的荧光探针(9,13)。该家族的一个成员,时间蛋白D,对红细胞有溶解作用(13)。最近对两种不同的颞蛋白B对映体在粪便环境中的稳定性进行的研究表明,肽的D异构体的失活速度比L异构体慢(14)。因此,当考虑治疗用途时,D异构体可能更可取。
颞蛋白与脂质的相互作用尚未得到详细研究。为了更深入地了解这些肽的作用模式,并将其与我们实验室先前研究过的其他两种抗菌肽(15)马盖宁2和吲哚肽进行比较,我们研究了颞蛋白B(LLPIVGNLLKSLL-NH2)和颞蛋白L(FVQWFSKFLGRIL-NH2)与模型膜的相互作用。更具体地说,我们研究了它们插入脂质单层的能力,使用二苯基己三烯(DPH)1和芘标记的磷脂对脂质双层结构动力学的影响,以及对巨大脂质体形态的影响。
材料和方法
材料。Hepes和EDTA来自Sigma。1-硬脂酰-2-油酰基-sn-甘油-3-磷酸胆碱(SOPC)、1-棕榈酰-2-油酰基-sn-甘油-3-磷酸甘油(POPG)和–胆固醇来自Avanti极性脂质(Alabaster,AL)。荧光磷脂类似物1-棕榈酰-2-[10-(芘-1-基)癸酰基]-sn-甘油-3-磷酸胆碱(PPDPC)来自K&V Bioware(芬兰埃斯波)和EGA Chemie(德国斯坦海姆)的二苯基己三烯(DPH)。使用高精度电子天平(Cahn、Cerritos、CA)通过重量法测定未标记脂质的浓度,并通过341 nm处的吸光度,分别使用38000和88000 cm-1的摩尔消光系数测定含芘磷脂和DPH的浓度。在用氯仿/甲醇/水(65:25:4,v/v/v)开发的硅酸涂层板(德国达姆施塔特默克公司)上,通过薄层色谱法检查脂质的纯度。碘染色后以及在适当情况下,紫外线照射后对平板进行检查,未发现任何杂质。合成颞叶素从塔纳实验室(德克萨斯州休斯顿)购买。通过HPLC分析肽的纯度(颞蛋白B和颞蛋白L的纯度分别大于90%和大于94%),并通过自动Edman降解和质谱验证其序列。通过重量法和定量离子交换柱色谱法以及茚三酮衍生法测定肽浓度。
肽对脂质单层的渗透。使用磁力搅拌的圆形聚四氟乙烯孔(多孔板,亚相体积1.2 mL,Kibron Inc.,芬兰赫尔辛基)测量肽插入位于空气/水界面上的脂质单层。表面压力(π)通过连接到连接到奔腾PC的微量天平(DeltaPi,Kibron Inc.)上的Wilhelmy线进行监测。所示脂质以氯仿(约1 mg/mL)的形式扩散到空气/缓冲液(5 mM Hepes,0.1 mM EDTA,pH 7.0)界面上。随后,在将temporins(0.3µM最终浓度)注入亚相之前,允许脂质单层在不同初始表面压力(π0)下沉淀约15分钟。注射肽后π的增量在大约30分钟内完成,初始表面压力(π0)与肽渗透到膜中后观察到的值之间的差值取为∆π.数据表示为∆πvsπ0。这些图还得出了与阻止肽插入膜中的脂质侧向堆积密度相对应的临界表面压力πc。所有测量均在环境温度下进行,一式三份(≈+24°C)。
制备大的单层囊泡(LUV)。将适量的脂质储备溶液混合在氯仿中以获得所需的组合物,其中PPDPC(X)0.01)或DPH(X)0.002)包括作为荧光探针。在氮气流下去除溶剂,随后在减压下将脂质残留物保持至少2小时。然后在50°C下将干脂质在5 mM Hepes、0.1 mM EDTA、pH 7.0中水合,以产生1 mM的脂质浓度。使用Liposofast低压均质器(加拿大渥太华Avestin)通过两个聚碳酸酯过滤器(孔径100 nm,Bedford,MA)堆叠挤出所得分散体,以获得平均直径在111和117 nm之间的大单层囊泡(17)。
Ie/Im的测量。芘的辐照≈344 nm产生单体激发态,该激发态通过发射最大值为的光子松弛回到基态≈398 nm(Im),精确的峰能量和光谱结构取决于溶剂极性。如果芘的局部浓度足够高,激发单体可能与基态芘碰撞,形成激发二聚体(准分子)。准分子通过发射以中心为中心的宽而无特征的量子带,解离回两个基态芘≈480纳米(18,19)。用Perkin Elmer LS50B荧光光谱仪和磁力搅拌恒温反应杯室测量用PPDPC(X)0.01)标记的LUV的荧光发射光谱。激发和发射均使用4 nm的带宽。使用的脂质浓度为20µM,温度保持在25°C。添加适量肽后,在记录光谱之前,样品平衡5分钟。取三次扫描的平均值,并分别测量Im和Ie处的发射强度。≈取398和480 nm进行
DPH的荧光各向异性(r)。DPH在X)0.002处包含在脂质体中。使用的脂质浓度为20µM,温度保持在25°C。使用带有Perkin Elmer LS50B的宝丽来膜式过滤器以L格式测量极化发射。使用10 nm带宽,在360 nm激发和450 nm发射下测量DPH的荧光各向异性r,并使用Perkin Elmer提供的软件例程计算其值。
形成巨大脂质体。如其他地方所述(20-22)制备了巨大脂质体。将溶解在二乙醚/甲醇(9:1,v/v,浓度为1 mM)中的约1-3µL指示脂质摊铺在两个铂电极的表面上,然后在氮气流下干燥。通过在真空中抽空1小时来去除可能的有机溶剂残留。在蔡司IM-35倒置荧光显微镜的工作台上放置一个带有连接电极和石英窗口底部的玻璃室。在添加1.3 mL pH值为7.4的0.5 mM Hepes缓冲液之前,施加交流电场(频率为4 Hz、振幅为0.2 V的正弦波函数)。在水合作用的第一分钟内,电压增加到1.0 V。2小时后关闭交流电场,使用霍夫曼调制对比度(HMC)光学系统观察到巨大的脂质体,其物镜为10×/0.25(modulation optics Inc.,纽约)。巨大脂质体的大小是通过微管的运动校准图像,将其作为微操作器步长(50 nm)的适当倍数(带有MC2000控制器的MX831、SD仪器、Grants Pass或)。使用珀尔贴冷却的12位数字CCD摄像机(C4742-95,日本滨松)与计算机连接,并通过摄像机制造商提供的软件(HiPic 5.0.1)进行操作,记录图像。
微量注射技术。内部尖端二聚体>0.5µm(23)的微移液管由硼硅酸盐毛细管(外径1.2 mm)通过微处理器控制的水平拉拔器(P-87,Sutter Instrument Co.,Novato,CA)制成。将指示量的肽溶液(10 mM Hepes中的0.5 mM,0.1 mM EDTA,pH 7.0)施加到单个巨大脂质体的外表面,作为一系列约20 fL的单次注射,用气动微注射器(PLI-100,Medical Systems Corp.,Greenvale,NY)输送。为了便于操作,只使用附着在电极表面的囊泡。所有实验均在环境温度下进行(≈+并重复至少10次。
结果
颞叶素对脂质单层的渗透。在将配体(如肽)注射到水/气界面上的脂质单分子膜中后,将其插入到水/气界面上的脂质单分子膜中会导致增加∆π、在表面压力下。获得的数据表示为∆π、作为薄膜初始表面压力的函数,π0(15,16)。表面压力变化的幅度可用于评估肽-脂质相互作用的相对能量,并表征肽的脂质特异性(综述见16)。先前采用该技术的研究表明,抗菌肽,如马盖宁2和吲哚肽,可以有效插入脂质膜(15)。同样,颞蛋白B和颞蛋白L均有效插入脂质单层,如初始压力(π0)为0.20)时的SOPC/POPG膜(XPOPG)所示≈15 mN/m(图1)。然而,两种肽表面压力增加的动力学明显不同。因此,在亚相中加入颞蛋白B(图1,图A)后,π出现快速瞬态峰值,然后是松弛,然后是π缓慢增加。这些动力学与脂质成分无关,初始表面压力在12-36 mN/m范围内变化。相反,对于Temporan L,表面压力值以连续方式增加,在大约40分钟内达到平台(图1,面板B)。现阶段未对动力学进行更详细的研究。
随后从以下方面分析了来自上述类似测量的数据:∆πvsπ0(图1,面板C和D)。时间蛋白B很容易插入到高达π0的SOPC单分子膜中≈39 mN/m,其中∆π-π0数据点外推到∆π)0.因此,该值表示该肽不再插入SOPC单层的临界包装压力πc(图1,面板c)。与其净正电荷+2一致,当膜中酸性磷脂POPG的含量增加时,时间蛋白B向脂质单层的渗透逐渐增强(图1,面板C),πC的值急剧增加。有趣的是,当XPOPG在0.10和0.40之间变化时∆temporin B的πvsπ0数据在表面压力为的情况下显示出一个“交叉”点≈32 mN/m。具体来说,在后一个表面压力以下,增加∆π随着膜中POPG的增加而增加,而在π0>32mn/m时观察到相反的情况。πc的值在序列中降低≈64,≈55,和≈当XPOPG分别从0.10增加到0.20和0.40时,为48 mN/m(图1,面板E)。将胆固醇(X)0.10)包含在SOPC膜中抑制了该肽在π0处插入单层≈27 mN/m(图1,面板C),而πC增加到≈4200万/米。
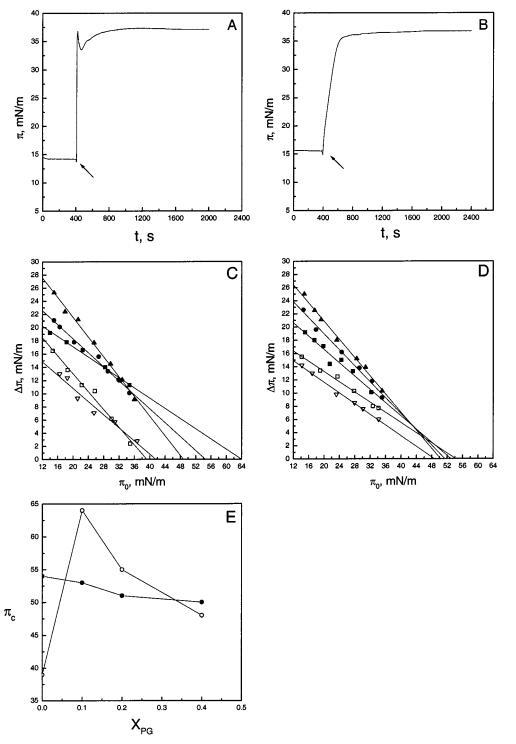
图1:将时间蛋白B和L插入脂质单层。在初始表面压力(π0)分别为14.6和15.5 mN/m的情况下,在SOPC/POPG(XPOPG)0.2)单层下方注射颞蛋白B(面板a)或颞蛋白L(面板B)后,表面压力随时间的增加而增加。肽(0.3µM最终浓度)的添加用箭头标记。表面压力增量(∆π)将0.3µM颞蛋白B(图C)或颞蛋白L(图D)添加到亚相中导致的脂质单分子膜的数量表示为初始表面压力(π0)的函数。SOPC中POPG(XPOPG)的含量分别为0(0)、0.10(9)、0.20(b)和0.40(2)。还显示了将肽插入具有Xchol)0.1(3)的SOPC膜中。图E显示了πc的值,分别作为渗透时间蛋白B(O)和时间蛋白L(B)时XPOPG的函数。每个数据点代表三次测量的平均值。标准偏差在0.1和0.8 mN/m之间变化,为清晰起见,未显示。
也观察到由于颞蛋白L,π显着增加,表明其渗透到SOPC单层(图1,面板D)。然而,temporin L明显比temporin B更具膜活性,具有πc≈54 mN/m。与temporin B类似,单分子层中POPG含量的增加增强了temporin L的渗透性,增加了∆π以下≈43 mN/m(图1,面板D)。然而,在表面压力的后一个“交叉”值以上,由于时间蛋白L引起的π增量被POPG略微衰减,导致在存在POPG的情况下πc减小,转移到≈53,≈51,和≈XPOPG时为50 mN/m),分别为0.10、0.20和0.40(图1,面板E)。至于颞蛋白B,由颞蛋白L引起的π增量被胆固醇(Xchol)0.10减弱,而πc从≈54至≈48 mN/m(图1,面板D)。
颞蛋白对双层脂质动力学的影响。随后通过记录掺入LUVs的芘标记荧光磷脂类似物PPDPC(X)0.01)的发射光谱来研究肽与脂质体结合的后果。对于含有脂质类似物(如PPDPC)的单个芘部分,稳态Ie/Im值反映了膜中荧光团的横向迁移率和局部浓度(18,19)。然而,我们之前的研究比较了两种其他抗菌肽(马盖宁2和吲哚肽)对膜脂质动力学的影响,发现这些肽也影响芘标记脂质的量子产率(15)。对于SOPC LUV,添加temporin B分别导致芘单体和准分子发射Im和Ie的显着减少,最大减少约40%(图2,面板a和B)。芘荧光带的猝灭是双相的,当时间蛋白B与脂质的摩尔比约为1:12时,量子产率显着降低(图2,面板a和B)。在胆固醇(Xchol)0.10)存在的情况下,temporin B对芘单体和准分子荧光的猝灭也很明显,尽管发射的减少量小于在SOPC LUV中观察到的减少量(图2,面板A和B)。然而,当酸性磷脂POPG存在于XPOPG)0.10时,Im中只有微不足道的变化是明显的,当XPOPG从0.20增加到0.40时,时间蛋白B导致Im以渐进的方式增加,Im的初始增量在大约2/1的POPG/时间蛋白B化学计量比下趋于稳定。在达到饱和响应之前,Im中线性增量的斜率随着XPOPG而增加(图2,面板C)。在POPG存在的情况下,颞蛋白B也增加了Ie。在XPOPG)0.40时,Ie的值逐渐增加,直到颞蛋白B:脂质摩尔比为≈1:15,对应于≈1/6,而当超过该化学计量比时,Ie降低。图3,面板A中显示了由于颞蛋白B引起的Ie/Im的变化。具体而言,SOPC LUV的Ie/Im增加了1.1倍,而在胆固醇(Xchol)0.10的存在下,观察到Ie/Im的微小变化,面板A)。将XPOPG从0.10增加到0.20会逐渐增强Ie/Im的增量。有趣的是,在XPOPG)0.40时,当时间蛋白B:脂质摩尔比为≈1:30.然而,在这一肽:脂比之上,颞叶素B导致Ie/Im降低,而在这一肽:脂比之上,Ie/Im的变化不显着≈1:6.
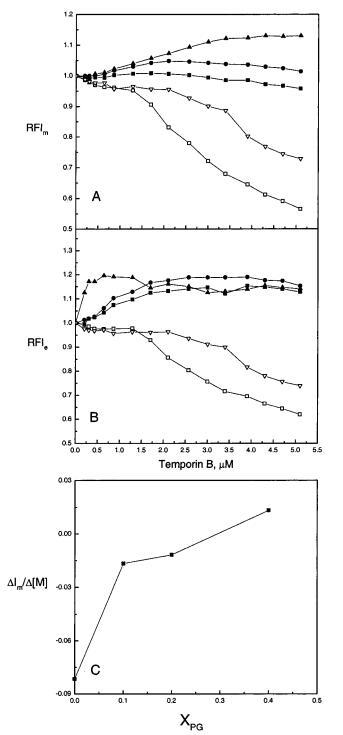
图2:通过芘标记的磷脂PPDPC(X)0.01的荧光评估颞叶素B对LUV中脂质动力学的影响,分别显示为芘单体和准分子发射强度Im(图A),Ie(图B)的变化,以及在RFIm中达到饱和反应之前Im增加的斜率(图C)。脂质体由SOPC组成,XPOPG为0(0)、0.10(9)、0.20(b)和0.40(2),Xchol为0.10(3)。总体积为2 mL的5 mM Hepes,0.1 mM EDTA,pH 7.0,脂质浓度为20µM。使用循环水浴将温度保持在25°C。每个数据点代表三次测量的平均值。标准偏差小于0.02(面板A和B)和0.004(面板C),为清晰起见,未显示。
芘标记的脂质PPDPC的Ie/Im增强通常被评估为是由于探针的侧向分离或脂质侧向扩散速率增加所致。为了解决这两种相互非排斥机制之间的矛盾,我们测量了膜中DPH的稳态荧光各向异性r(图3,面板B)。DPH是一种小的、疏水的杆状荧光团,其中很大一部分位于双层的碳氢化合物区域,平行于膜脂的长轴。其发射各向异性可用于评估酰基链顺序(有关综述,请参阅24)。增加脂质堆积后,观察到酰基链顺序增加,从而减少脂质侧向扩散(25,26)。颞叶素B对SOPC和SOPC/胆固醇(Xchol)0.10)LUVs的r仅产生了微不足道的变化(图3,面板B)。因此,SOPC-luv的酰基链顺序没有变化以及Ie/Im的增加表明PPDPC很可能富集到微域中。随着XPOPG的增加,添加颞蛋白B后,脂质堆积和酰基链顺序逐渐增强。因此,膜横向扩散的增加不能导致观察到的Ie/Im和颞叶素B诱导的脂质分离的增加;即。,可以得出PPDPC在膜中聚集的结论。然而,当考虑淬火过程的影响时,解释变得更加复杂。
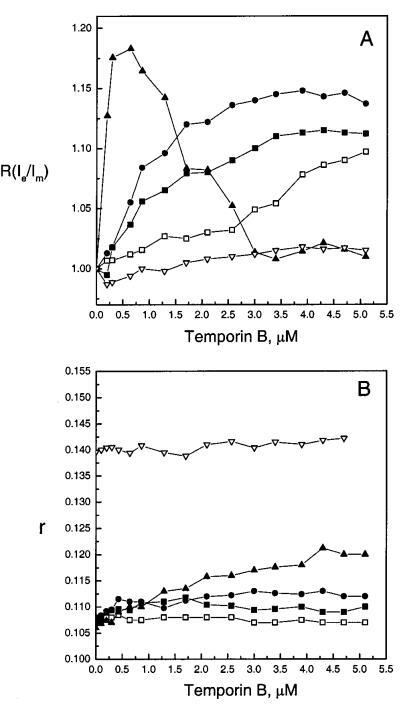
图3:通过芘标记的磷脂PPDPC(X)0.01)的荧光和DPH(X)0.002的稳态发射各向异性r评估颞叶素B对LUV中脂质动力学的影响,显示为具有XPOPG)0(0)、0.10(9)、0.20(B)和0.40(2)的SOPC脂质体中归一化准分子与单体比率r(Ie/Im)(图A)和各向异性r(图B)的变化,Xchol)0.10(3)。每个数据点代表三次测量的平均值。标准偏差小于0.02(面板A)和0.004(面板B),为清楚起见,未显示。否则情况如图2图例所示。
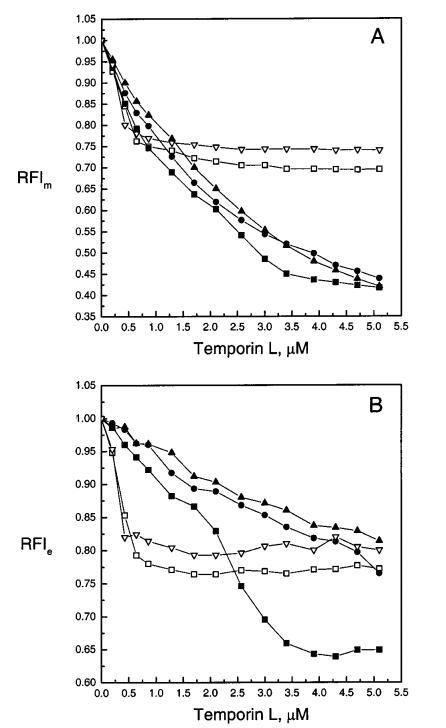
图4:通过芘标记的磷脂PPDPC(X)0.01的荧光评估颞叶素L对LUV中脂质动力学的影响,分别显示为芘单体和准分子发射强度Im(图A)和Ie(图B)的变化。脂质体由SOPC组成,XPOPG为0(0)、0.10(9)、0.20(b)和0.40(2),Xchol为0.10(3)。每个数据点代表三次测量的平均值。标准偏差小于0.02,为清晰起见,未显示。否则情况如图2图例所示。