




合作客户/
拜耳公司 |
同济大学 |
联合大学 |
美国保洁 |
美国强生 |
瑞士罗氏 |






相关新闻Info
推荐新闻Info
-
> 芬兰Kibron全系列表面张力仪核心测量原理是什么
> 芬兰Kibron表面张力仪 vs 主流品牌表面张力仪对比分析
> 6∶2氟调磺酸在饱和与非饱和土壤中的迁移行为研究
> 6∶2氟调磺酸表面活性及其在气-水界面的吸附特性
> 基于表面张力测定量化6∶2氟调磺酸表面活性
> 不同类型表面活性剂对氨气-水溶液界面行为的调控规律研究
> 阶梯内嵌式复合层线路板的涂覆控制装置及其控制方法——基于双组分表面张力驱动与共形锁定的技术分析
> LB膜分析仪驱动的界面功能化技术革新:FAST-LAM的性能对标研究
> 面向高温高盐油藏的内烯烃磺酸盐驱油表面活性剂性能研究
> 离子类型与浓度对内烯烃磺酸盐降低油水界面张力的协同效应
细胞表面PH对于生物多肽生物活化的潜在重要性——结果和讨论、结论、致谢!
来源: 浏览 2692 次 发布时间:2022-01-04
结果和讨论
细胞和探针肽的选择
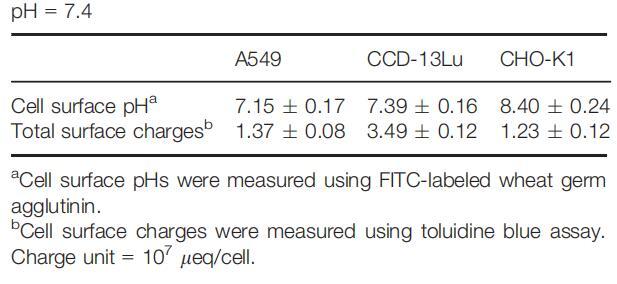
虽然细胞外环境通过影响细胞表面和整体溶液之间的质子交换而影响细胞表面pH值,但在pH值固定的生理条件下,细胞表面pH值将由细胞的化学和生物学特性决定,包括细胞的质子生产能力、细胞大小、细胞的大小、细胞的大小、细胞的大小、细胞的大小、细胞的大小、细胞的大小、细胞的大小、细胞的大小以及细胞的大小,以及细胞表面的pH缓冲能力。因此,在生理条件下,组织细胞可能具有特定的甚至可区分的表面pHs。我们测量了一组来自不同器官/组织的细胞在生理pH下的细胞表面pH值,发现所有受试细胞都具有不同的表面pH值(表1)。
表1:生理条件下测得的细胞表面pH值和电荷
裂解肽是一组阳离子肽,主要作用于细胞膜,通过引起细胞裂解杀死细胞。对裂解肽与细胞相互作用的研究表明,肽向结合平面的转变和插入膜是裂解肽诱导细胞膜裂解的关键(6,9,11)。我们选择具有pH依赖性细胞裂解活性的裂解肽作为探针,研究细胞表面pH对肽-细胞相互作用的潜在影响。如果pH敏感的裂解肽能够感应到本体溶液和细胞表面之间的pH差,从而改变肽-细胞相互作用并显示出改变的细胞裂解活性,则可以评估细胞表面pHs的潜在药学意义。
我们已经开发了一种构建具有pH依赖性细胞裂解活性的裂解肽的通用方法(9,12)。一些pH敏感的裂解肽表现出高达30倍的活性增加,以响应降低的培养基pH(12)。由于肽对细胞膜的静电吸引是将肽带到细胞表面的第一步,也是关键步骤,因此肽上的净电荷将对肽与细胞的相互作用产生显着影响。在这方面,仅选择具有相同(+1)电荷的pH敏感裂解肽(表2)。除了电荷外,肽的二级结构还可能显着影响肽与细胞的相互作用。肽链折叠成特定构象(通常为α螺旋结构)证明是肽插入细胞膜的必要且能量有利的过程(6,13)。在本研究中,选择了具有相同二级结构(a-螺旋)的pH敏感裂解肽。这种肽和细胞选择使我们能够直接比较从不同细胞和肽获得的结果。
表2:所选裂解肽的性质
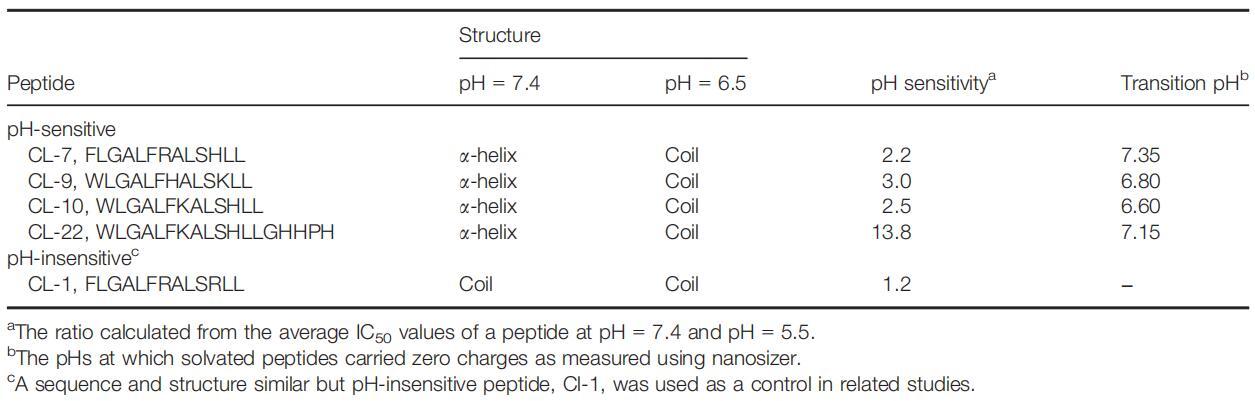
肽CL-1是一种pH不敏感的裂解肽(表2)。尽管其序列和结构与其他pH敏感肽相似,但其细胞裂解活性几乎不受pH的影响。本研究以肽CL-1作为阴性对照。
pH敏感裂解肽对不同表面pH值细胞的活性
首先在CHO-K1细胞上测试pH敏感裂解肽的活性,测得的细胞表面pH=8.40。由于CHO-K1的表面pH值远高于所有选定肽的过渡pH值(表2),因此所有肽在从散装溶液转移到细胞表面时都会携带相同的负电荷(pH=7.4)。四种测试的pH敏感裂解肽在CHO-K1细胞上表现出较低且几乎相同的细胞裂解能力(图2)。
图2:在CHO-K1细胞上测试的pH敏感裂解肽(40 lM)的细胞裂解活性。研究在无血清培养基(pH=7.4)中进行。对照组未添加肽。对照空白对照组和100%裂解对照组,将细胞裂解(乳酸脱氢酶释放)标准化为细胞死亡百分比。数据代表至少三个独立测试的平均值和标准差。
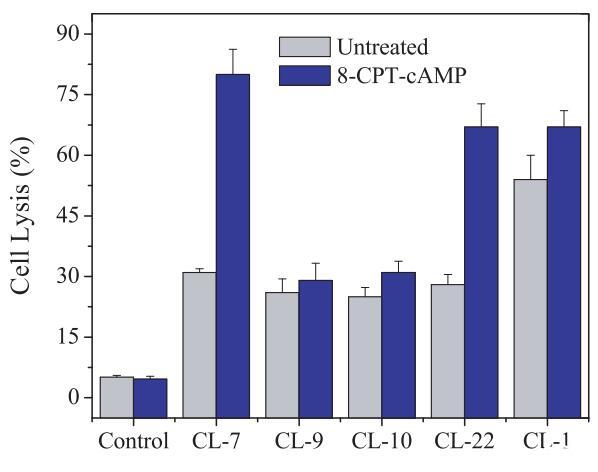
为了研究细胞表面pH对pH敏感裂解肽与细胞相互作用的潜在影响,我们使用质子通道调节剂8-CPT-cAMP(14)调节细胞表面pH。在正常细胞培养条件下,8-CPTcAMP对CHO-K1细胞的活力没有影响(图2)。然而,通过8-CPT-cAMP,CHO-K1的细胞表面pH值迅速(在30分钟内)从pH=8.4降至pH=6.90,而培养基pH值保持不变,保持在pH=7.4(表3)。尽管8-CPT-cAMP处理的CHO-K1细胞的表面pH值仍高于肽CL-9(pH=6.80)和肽CL-10(pH=6.60)的过渡pH值,但低于肽CL-7(pH=7.35)和肽CL-22(pH=7.15)的过渡pH值。有趣的是,四种pH敏感裂解肽对8-CPT-cAMP处理的CHO-K1细胞表现出不同的细胞裂解能力:肽CL-9和CL-10保持与未处理的CHO-K1细胞相同的活性,而肽CL-7和CL-22显示出显着增加(约两倍)的细胞裂解活性(图2)。肽的活性结果与未处理和8-CPT-CAMP处理的CHO-K1细胞的表面pH值相匹配,表明细胞表面pH值参与pH敏感肽的激活。
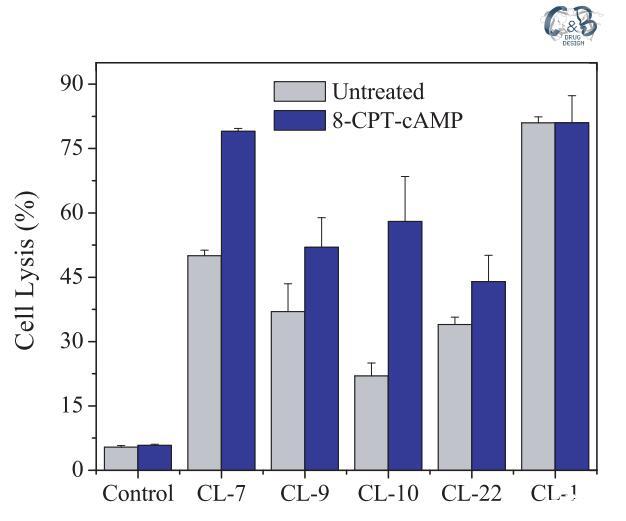
为了排除上述结果由CHO-K1细胞的某些固有特性引起的可能性,我们在另一个选定的细胞A549上进行了相同的实验,其细胞表面电荷与CHOK1细胞几乎相同(表1)。由于肽将受到来自A549和CHO-K1细胞的相同拉力,静电吸引对肽-细胞相互作用的潜在影响被最小化或消除。A549细胞的表面pH值为7.15,高于肽CL-9、CL-10和CL-22的过渡pH值,但低于肽CL-7的过渡pH值(pH值为7.35)。同样,CL-7被证明是A549细胞上最活跃的肽(图3)。在存在8-CPT-cAMP的情况下,A549细胞的表面pH值降低至pH=6.55(表3),低于所有受试裂解肽的过渡pH值(表1)。因此,四种pH敏感裂解肽对8-CPTcAMP处理的A549细胞的裂解活性显着增加(图3)。应注意的是,与pH敏感肽不同,当细胞被8-CPT-cAMP处理时,pH不敏感肽CL-1对A549和CHO-K1细胞的活性几乎没有变化(图2和图3)。
图3:在A549细胞上测试的pH敏感裂解肽(40 lM)的细胞裂解活性。研究在无血清培养基(pH=7.4)中进行。对照组未添加肽。对照空白对照组和100%裂解对照组,将细胞裂解(乳酸脱氢酶释放)标准化为细胞死亡百分比。数据代表至少三个独立测试的平均值和标准差。
细胞表面pHs影响肽活化的进一步证据来自使用另一种质子通道调节剂巴丝霉素A1(15)的类似研究。与8-CPT-cAMP一样,巴非霉素A1对细胞活力的影响非常有限(图4),但巴非霉素A1处理导致细胞表面pHs增加。此外,巴非霉素A1能够逆转8-CPT-cAMP对细胞的影响,并使细胞表面pHs恢复到正常水平或更高水平(表3)。研究表明,最初在8-CPT-cAMP处理的细胞上观察到的肽的细胞裂解活性的升高被逆转,因为细胞表面pH值的增加超过了巴丝霉素A1的肽的过渡pH值(图4)。
表3:8-CPT-cAMP-和巴非霉素A1在具有生理pH的培养基中诱导的细胞表面pH变化
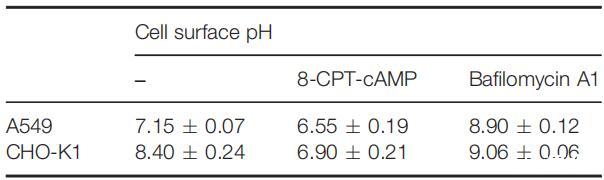
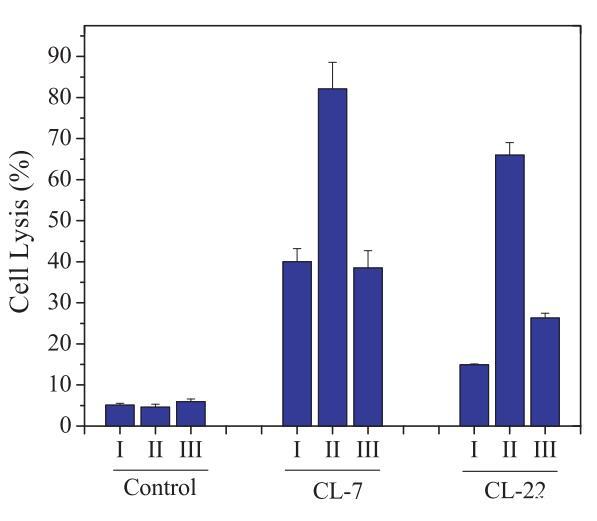
图4:在CHO-K1细胞(I)、用8-CPT-cAMP处理的CHO-K1细胞(II)和用8-CPT-cAMP处理的CHO-K1细胞上测试的细胞表面pHs调节PH敏感裂解肽CL-7和CL-22的细胞裂解活性,然后是巴丝霉素A1(III)。表3提供了不同处理对应的细胞表面pHs。在所有实验中,肽浓度固定在40 lM。对照组未添加肽。对照空白对照组和100%裂解对照组,将细胞裂解(乳酸脱氢酶释放)标准化为细胞死亡百分比。数据代表至少三个独立测试的平均值和标准差。
机理研究
肽与细胞膜的相互作用包括三个步骤:(i)阳离子肽对细胞表面的静电吸引;(ii)肽向结合平面的转变;(iii)将肽插入细胞膜。通过研究pH敏感肽CL-7诱导的膜损伤动力学,研究了肽-细胞相互作用的细节(图5A)。CL-7诱导的A549细胞膜渗漏呈时间依赖性,并呈线性曲线。尽管在8-CPT-cAMP处理的A549细胞上,CL-7介导的细胞膜损伤在开始时遵循相同的曲线,但它偏离了线性曲线,并在培养15-20分钟后加速(图5A)。在脂质单层膜上进行的平行膜张力研究表明,CL-7与细胞膜的相互作用以表面张力快速增加的初始阶段为特征,随后是表面张力松弛的第二阶段(图5B),与肽与膜结合和肽插入相关膜,分别。尽管CL-7与脂膜结合是一个快速过程(3-5分钟),但肽插入脂膜大约需要20分钟才能完成。这两个实验的结果非常吻合,这意味着细胞表面pH对CL-7的显着影响发生在肽-细胞相互作用的后期,即肽插入细胞膜。
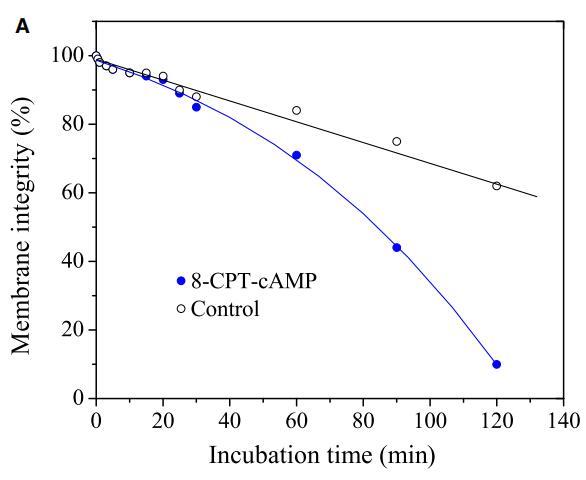
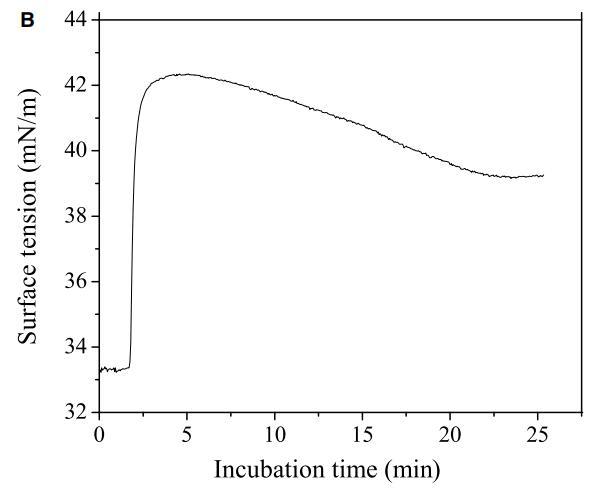
图5:(A)使用活/死试剂盒测定的肽CL-7诱导A549细胞膜损伤的动力学。在不同时间点添加肽溶液后拍摄荧光图像。计算绿色像素占总绿色和红色像素的百分比,以估计细胞膜的完整性。(B)肽CL-7结合和插入引起脂质单层的表面张力变化(DPPC/胆固醇/DPPS=50/10/2.5)。肽浓度保持在10 lM,脂质单层的初始表面张力设定为33 mN/m。
我们知道,由于肽上的净正电荷减少,含有组氨酸的pH敏感肽倾向于聚集,并且可以自组装成具有特定结构的肽聚集体(8,9,16)。在不同pH下聚集改变的肽是含组氨酸肽(6,9)pH依赖性活性的主要原因。使用1,8-ANS作为肽聚集体的疏水“口袋”探针的研究(图6A)表明,肽CL-7在生理pH下经历了自组装,如1,8-ANS在500 nm处的高发射强度和显着蓝移所示。使用SEM观察由纤维状超分子和纳米颗粒组成的CL-7聚集体(图6B)。然而,与其他含组氨酸的肽一样,CL-7聚集体不稳定,在酸性条件下溶解,因为组氨酸残基的咪唑基被质子化,并且肽分子之间的分子间排斥作用恢复(图6A)。这种pH值控制的CL-7聚集和溶解通过在高浓度(80 lM)肽溶液中进行的颗粒分析得到证实(图6C)。与肽聚集体(6,9)相比,自由形式的肽具有更高的膜亲和力和膜插入能力,这解释了pH敏感裂解肽对具有酸性表面pHs的细胞的强烈活性(图2、3和4)。
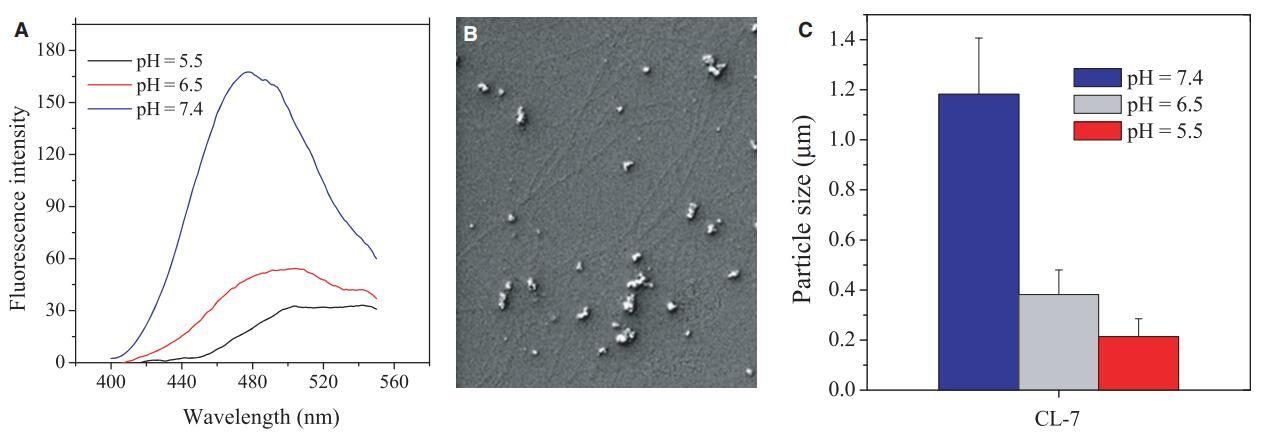
图6:(A)pH值影响不同pH值下溶液(40 lM)中CL-7的聚集。1-苯胺基萘-8-磺酸(1,8-ANS)浓度固定在20 lM,激发波长设置为369 nm。肽聚集溶解通过1,8-ANS发射峰在500 nm处的红移和荧光强度降低来指示;(B)pH=7.4的40 lM肽溶液中形成的CL-7聚集体的扫描电子显微镜图像;(C)隔夜培养后测量溶液中肽CL-7(80 lM)的粒径。数据代表至少三个独立测试的平均值和标准差。
除了肽聚集状态外,细胞表面pH值也可能影响pH敏感肽的二级结构。已经发现,肽插入细胞膜通常伴随着肽二级结构的变化(17)。肽链折叠成特定构象,通常为α螺旋结构,这被证明是肽插入细胞膜的必要过程(6)。尽管所有选定的pH敏感裂解肽在生理pH下均采用螺旋结构,但当环境pH降至肽的过渡pH以下时,它们变成随机线圈(表2和图7)。从无规螺旋转变为螺旋是肽插入膜的能量有利过程(6)。因此,细胞表面pH值也可能通过改变pH敏感肽的二级结构而影响其活性。
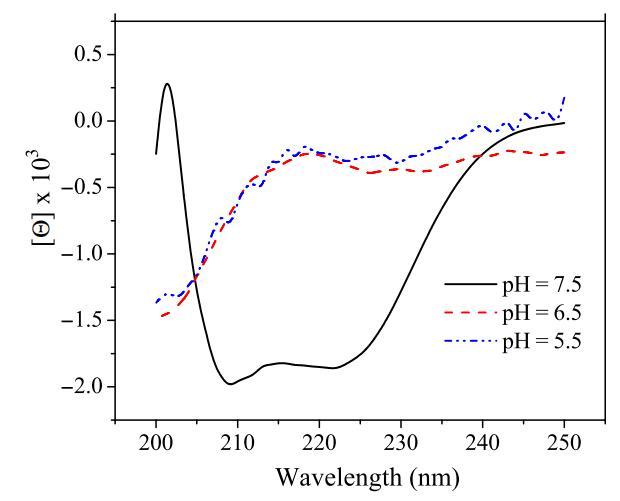
图7:pHs影响肽CL-7在20mM NaAc溶液中测得的圆二色谱光谱变化。以1.0 nm的间隔从250至190 nm收集数据,每个波长的积分时间为2秒。对五到十次扫描进行平均、平滑、背景减去,并转换为平均残余摩尔椭圆度(h)[度数/(cm2 dmol?1)]。
pH敏感肽对肿瘤的选择性
细胞表面pHs参与肽活化为设计具有所需细胞选择性的治疗性肽提供了新的机会。我们知道癌细胞通常具有较高的新陈代谢和较高的细胞表面缓冲能力(18)。因此,与正常细胞相比,癌细胞可能具有更多的酸性表面pHs。在我们研究中包括的正常-癌细胞对CCD-13Lu和A549之间,人肺癌细胞A549的表面pH值(pH=7.15)比正常人肺癌细胞CCD-13Lu(pH=7.4)的表面pH值(表1)高。尽管表面pH值差异如此之小,但它确实为我们提供了一个机会来测试利用细胞表面pH值来改善癌症治疗的可行性。
在细胞毒性研究中,选择了研究充分的pH敏感裂解肽PTP-7b(FLGALFKALSHL)(6,9)。由于PTP-7b的过渡pH(pH=7.25)低于A549的表面pH,但高于CCD-13Lu,因此裂解肽PTP-7b对A549的毒性可能比对CCD-13Lu细胞的毒性更大。选择PTP-7b的亲本肽PTP-7(FLGALFKALSKLL),一种pH不敏感的裂解肽(6,9)作为对照。如图8所示,尽管肽PTP-7对两种细胞的活性相同,但pH敏感肽PTP-7b对A549的毒性大约是对CCD-13Lu的三倍。
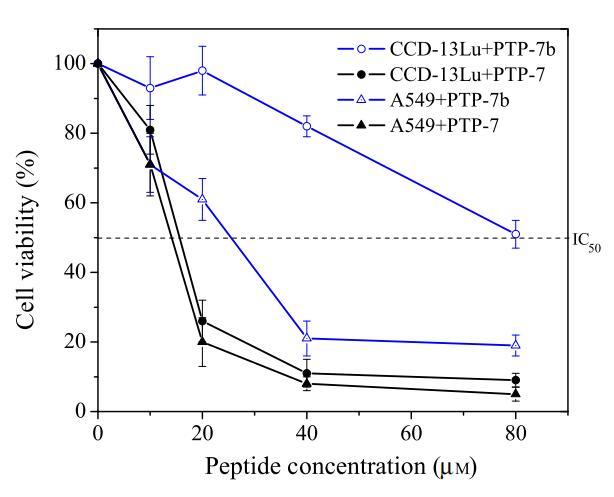
图8:pH敏感肽PTP-7b(FLGALFKALSHLL)对正常-癌细胞对CCD-13Lu和A549细胞的细胞毒性(MTT试验)。以pH不敏感肽PTP-7(FLGALFKALSKLL)作为对照。数据代表至少三个独立测试的平均值和标准差。
我们知道,与蛋白质相比,肽具有一些独特和优越的特性(19)。生物活性肽对生物体的功能和条件有积极的影响,并显示出对人类健康有益的特性。在过去的十年中,肽的研究得到了迅速的发展,而且这种发展很可能会继续下去。独特和特异的细胞表面pH值可能是设计具有所需细胞特异性和选择性的药物和治疗肽的新靶点。
致谢
这项工作得到了美国国立卫生研究院GM081874拨款的支持。陈先生是创新创业博士研究生奖学金的获得者。